
The Clinical Relevance of the EPH/Ephrin Signaling Pathway in Pediatric Solid and Hematologic Malignancies
 , , and
, , and
Abstract
:1. Introduction
1.1. Challenges and Opportunities in Pediatric Neoplasms: A Comprehensive Overview
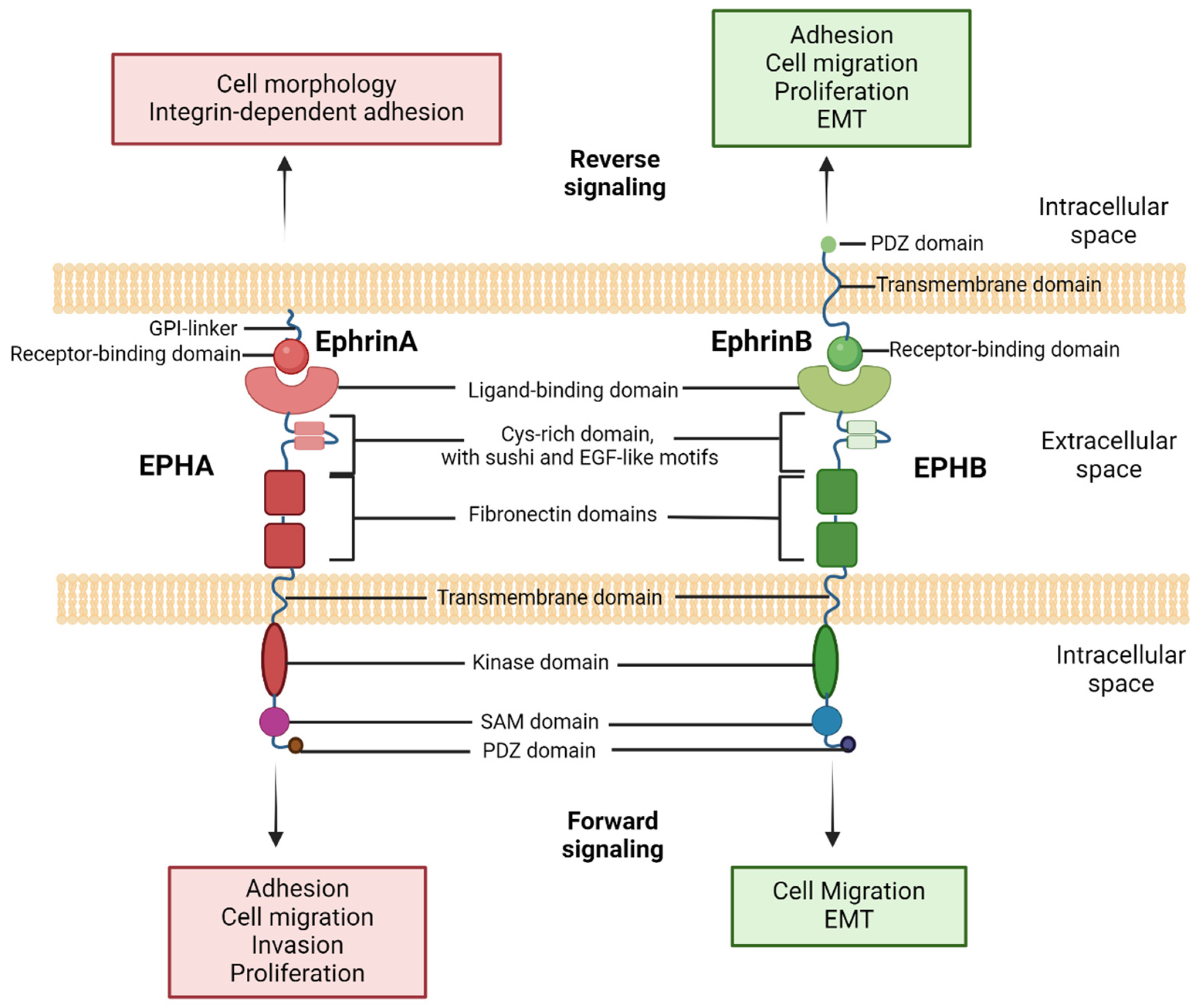
1.2. Insights into EPH/Ephrin Cellular Signaling
2. EPH/Ephrin Signaling in Physiologic Processes and Tumorigenesis
3. Exploring EPH/Ephrin Signaling in Solid Tumors of Pediatric Patients
3.1. Brain Tumors
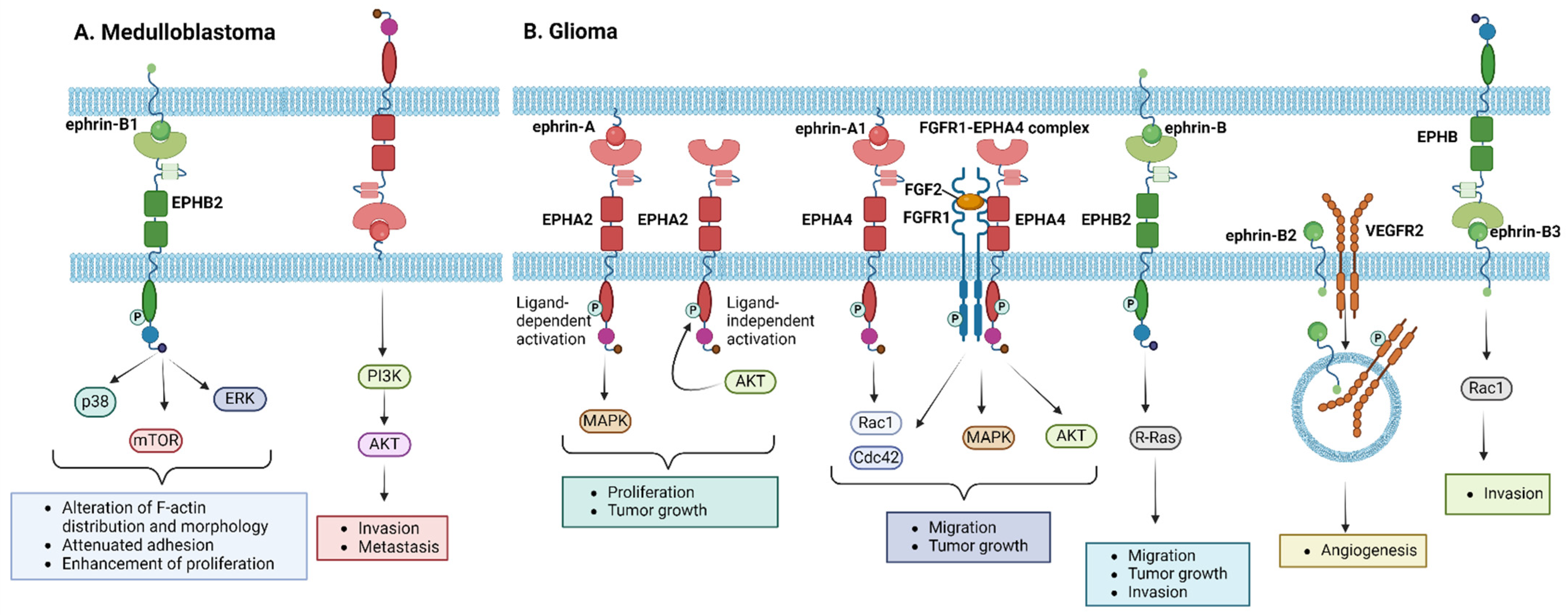
3.1.1. Medulloblastoma
3.1.2. Glioma
3.1.3. Ependymoma
3.1.4. Others
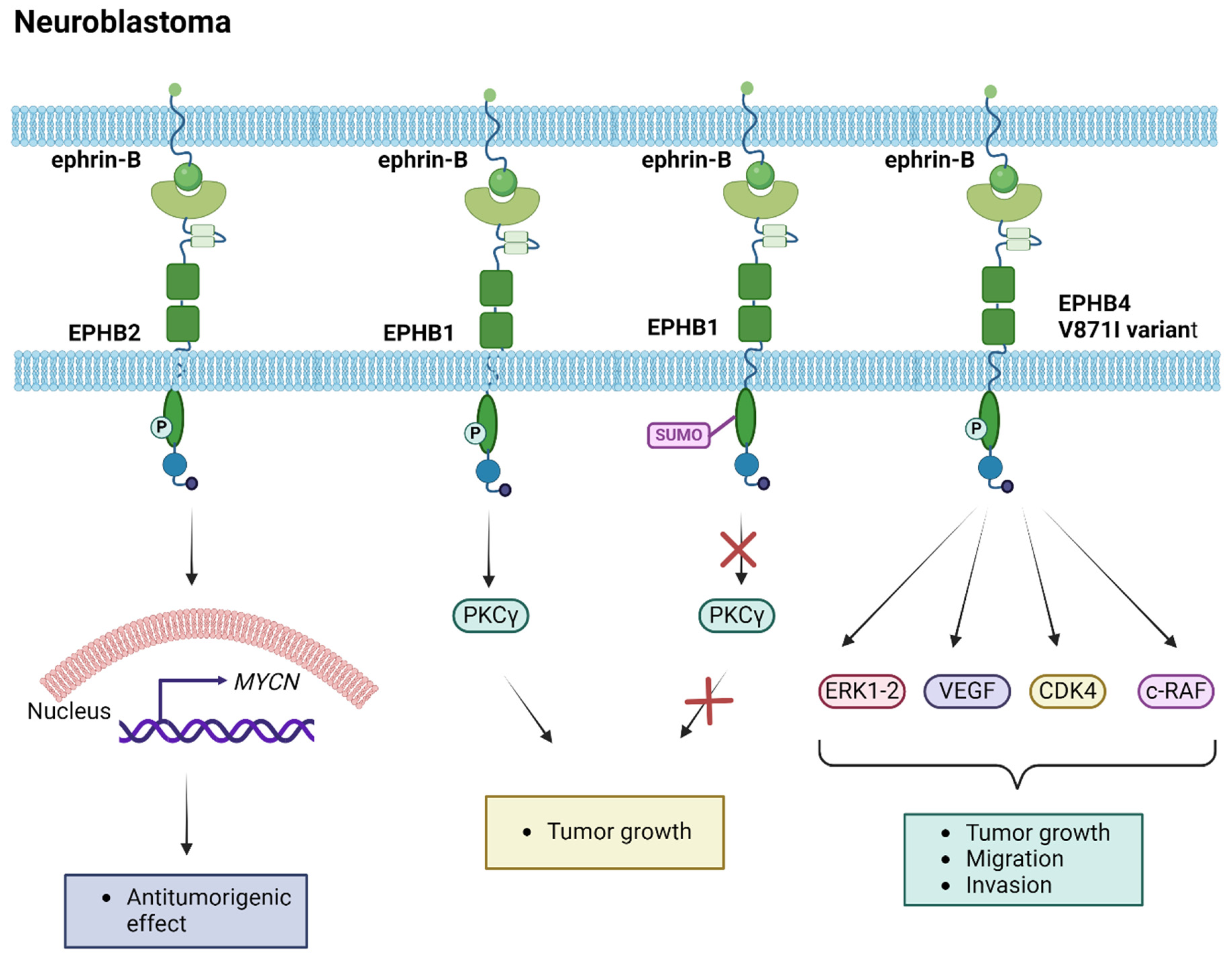
3.2. Neuroendocrine Tumors
3.3. Sarcomas
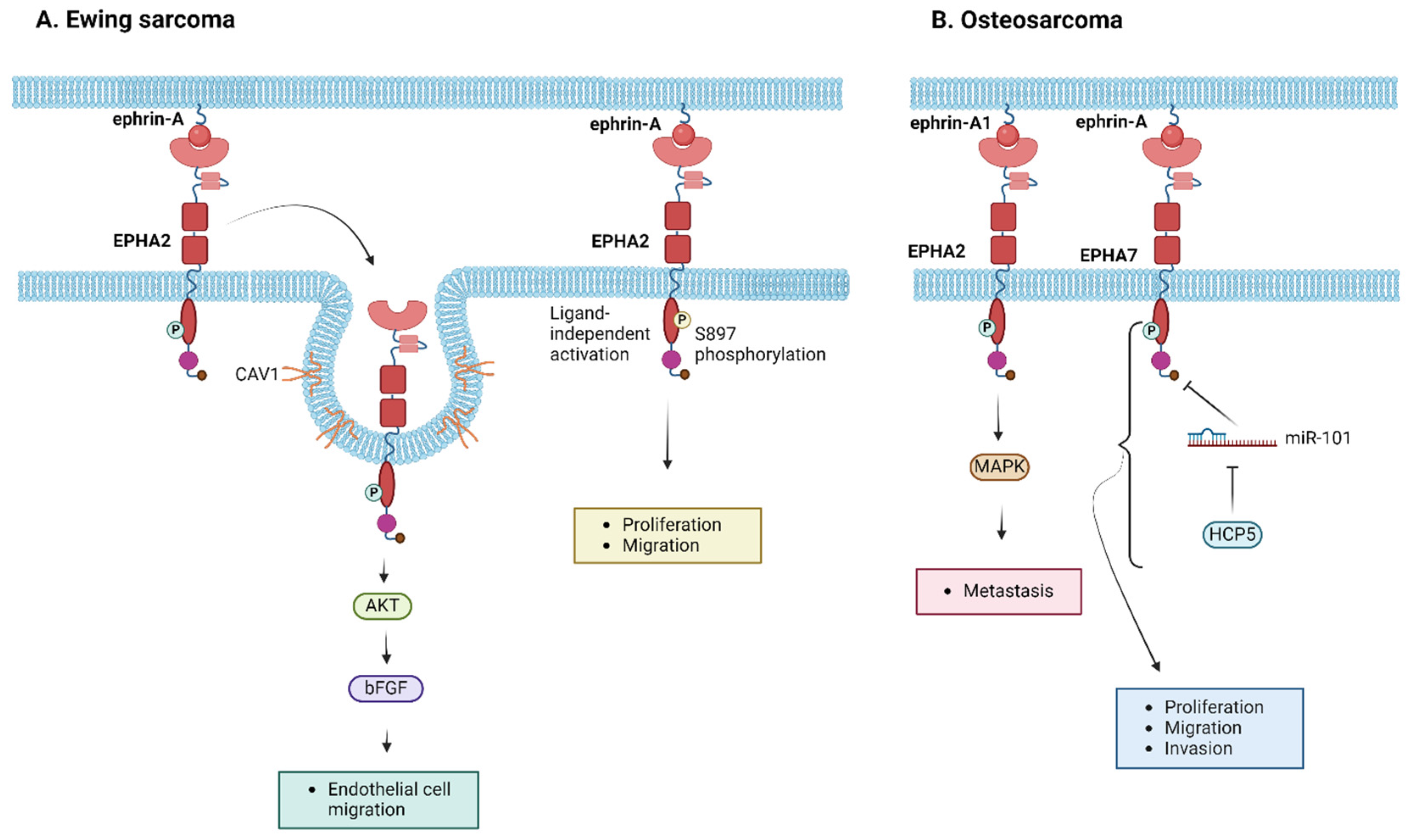
3.3.1. Ewing Sarcoma
3.3.2. Osteosarcoma
3.3.3. Rhabdomyosarcoma
3.4. Renal Tumors (Wilms)
4. EPH/Ephrin Signaling in Hematologic Malignancies of Pediatric Patients
5. Discussion
6. Limitations
7. Conclusions—Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- SIOP Europe. SIOP Europe Strategic Plan Update (2021–2026): A European Cancer Plan for Children and Adolescents. Available online: https://siope.eu/media/documents/siop-europes-strategic-plan-update-2021-2026.pdf (accessed on 20 February 2024).
- Namayandeh, S.M.; Khazaei, Z.; Najafi, M.L.; Goodarzi, E.; Moslem, A. GLOBAL Leukemia in children 0–14 statistics 2018, incidence and mortality and human development index (HDI): GLOBOCAN sources and methods. Asian Pac. J. Cancer Prev. 2020, 21, 1487–1494. [Google Scholar] [CrossRef] [PubMed]
- Udaka, Y.T.; Packer, R.J. Pediatric Brain Tumors. Neurol. Clin. 2018, 36, 533–556. [Google Scholar] [CrossRef] [PubMed]
- Terezakis, S.; Ladra, M. Pediatric rhabdomyosarcoma. Pediatr. Oncol. 2018, 20, 21–43. [Google Scholar]
- Laetsch, T.W.; Dubois, S.G.; Bender, J.G.; Macy, M.E.; Moreno, L. Opportunities and Challenges in Drug Development for Pediatric Cancers. Cancer Discov 2011, 11, 545–559. [Google Scholar] [CrossRef] [PubMed]
- Kern, S.E. Challenges in conducting clinical trials in children: Approaches for improving performance. Expert Rev. Clin. Pharmacol. 2009, 2, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Hirai, H.; Maru, Y.; Hagiwara, K.; Nishida, J.; Takaku, F. A Novel Putative Tyrosine Kinase Receptor Encoded by the eph Gene. Science 1987, 238, 1717–1720. [Google Scholar] [CrossRef] [PubMed]
- Lisabeth, E.M.; Falivelli, G.; Pasquale, E.B. Eph receptor signaling and ephrins. Cold Spring Harb. Perspect. Biol. 2013, 5, a009159. [Google Scholar] [CrossRef] [PubMed]
- Darling, T.K.; Lamb, T.J. Emerging roles for Eph receptors and ephrin ligands in immunity. Front. Immunol. 2019, 10, 454075. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, R.A.; Hunter, T. cDNA Cloning and Characterization of eck, an Epithelial Cell Receptor Protein-Tyrosine Kinase in the eph/elk Family of Protein Kinases. Mol. Cell. Biol. 1990, 10, 6316–6324. [Google Scholar]
- Zhou, R.; Copeland, T.D.; Kromer, L.F.; Schulz, N.T. Isolation and characterization of Bsk, a growth factor receptor-like tyrosine kinase associated with the limbic system. J. Neurosci. Res 1994, 37, 129–143. [Google Scholar] [CrossRef]
- Gilardi-Hebenstreit, P.; Nieto, M.A.; Frain, M.; Mattéi, M.G.; Chestier, A.; Wilkinson, D.G.; Charnay, P. An Eph-related receptor protein tyrosine kinase gene segmentally expressed in the developing mouse hindbrain. Oncogene 1993, 8, 1103. [Google Scholar] [PubMed]
- Maisonpierre, P.C.; Barrezueta, N.X.; Yancopoulos, G.D. Ehk-1 and Ehk-2: Two novel members of the Eph receptor-like tyrosine kinase family with distinctive structures and neuronal expression. Oncogene 1993, 8, 3277–3288. [Google Scholar] [PubMed]
- Aasheim, H.-C.; Patzke, S.; Hjorthaug, H.S.; Finne, E.F. Characterization of a novel Eph receptor tyrosine kinase, EphA10, expressed in testis. Biochim. Biophys. Acta—Gen. Subj. 2005, 1723, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Böhme, B.; Holtrich, U.; Wolf, G.; Luzius, H.; Grzeschik, K.H.; Strebhardt, K.; Rübsamen-Waigmann, H. PCR mediated detection of a new human receptor-tyrosine-kinase, HEK 2. Oncogene 1993, 8, 2857–2862. [Google Scholar] [PubMed]
- Andres, A.C.; Reid, H.H.; Zürcher, G.; Blaschke, R.J.; Albrecht, D.; Ziemiecki, A. Expression of two novel eph-related receptor protein tyrosine kinases in mammary gland development and carcinogenesis. Oncogene 1994, 9, 1461–1467. [Google Scholar] [PubMed]
- Boyd, A.W.; Ward, L.D.; Wicks, I.P.; Simpson, R.J.; Salvaris, E.; Wilks, A.; Welch, K.; Loudovaris, M.; Rockman, S.; Busmanis, I. Isolation and characterization of a novel receptor-type protein tyrosine kinase (hek) from a human pre-B cell line. J. Biol. Chem. 1992, 267, 3262–3267. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.; Watt, V.M. Eek and Erk, New Members of the Eph Subclass of Receptor Protein-Tyrosine Kinases. Oncogene 1991, 6, 1057–1061. [Google Scholar] [PubMed]
- Letwin, K.; Yee, S.-P.; Pawson, T. Novel protein-tyrosine kinase cDNAs related to fps/fes and eph cloned using anti-phosphorylation antibody. Oncogene 1988, 3, 621–627. [Google Scholar]
- Gurniak, C.B.; Berg, L.J. A new member of the Eph family of receptors that lacks protein tyrosine kinase activity. Oncogene 1996, 13, 777–786. [Google Scholar]
- Fox, G.M.; Holst, P.L.; Chute, H.T.; Lindberg, R.A.; Janssen, A.M.; Basu, R.; Welcher, A.A. cDNA cloning and tissue distribution of five human EPH-like receptor protein-tyrosine kinases. Oncogene 1995, 10, 897–905. [Google Scholar]
- Bartley, T.D.; Hunt, R.W.; Welcher, A.A.; Boyle, W.J.; Parker, V.P.; Lindberg, R.A.; Lu, H.S.; Colombero, A.M.; Elliott, R.L.; Guthrie, B.A. B61 Is a Ligand for the ECK Receptor Protein-Tyrosine Kinase. Nature 1994, 368, 558–560. [Google Scholar] [CrossRef] [PubMed]
- Wykosky, J.; Palma, E.; Gibo, D.M.; Ringler, S.; Turner, C.P.; Debinski, W. Soluble Monomeric EphrinA1 Is Released from Tumor Cells and Is a Functional Ligand for the EphA2 Receptor. Oncogene 2008, 27, 7260. [Google Scholar] [CrossRef] [PubMed]
- Takasugi, M.; Okada, R.; Takahashi, A.; Virya Chen, D.; Watanabe, S.; Hara, E. Small extracellular vesicles secreted from senescent cells promote cancer cell proliferation through EphA2. Nat. Commun. 2017, 8, 15729. [Google Scholar] [CrossRef] [PubMed]
- Dravis, C. Ephs, Ephrins and Bidirectional Signaling. Nat. Educ. 2010, 3, 22. [Google Scholar]
- Pasquale, E.B. Eph-Ephrin Bidirectional Signaling in Physiology and Disease. Cell 2008, 133, 38–52. [Google Scholar] [CrossRef] [PubMed]
- Georgakopoulos, A.; Litterst, C.; Ghersi, E.; Baki, L.; Xu, C.J.; Serban, G.; Robakis, N.K. Metalloproteinase/Presenilin1 Processing of EphrinB Regulates EphB-Induced Src Phosphorylation and Signaling. EMBO J. 2006, 25, 1242–1252. [Google Scholar] [CrossRef] [PubMed]
- Pasquale, E.B. Eph receptors and ephrins in cancer: Bidirectional signalling and beyond. Nat. Rev. Cancer 2010, 10, 165–180. [Google Scholar] [CrossRef] [PubMed]
- Arvanitis, D.; Davy, A. Eph/ephrin signaling: Networks. Genes Dev. 2008, 22, 416–429. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A.M.; Shi, J.; Wolf, A.M.; Lu, C.-C.; King, L.A.; Zou, Y. Wnt–Ryk signalling mediates medial–lateral retinotectal topographic mapping. Nature 2006, 439, 31–37. [Google Scholar] [CrossRef]
- Schlessinger, J. Receptor tyrosine kinases: Legacy of the first two decades. Cold Spring Harb. Perspect. Biol. 2014, 6, a008912. [Google Scholar] [CrossRef]
- Pasquale, E.B. Eph receptor signalling casts a wide net on cell behaviour. Nat. Rev. Mol. Cell Biol. 2005, 6, 462–475. [Google Scholar] [CrossRef] [PubMed]
- Kania, A.; Klein, R. Mechanisms of ephrin–Eph signalling in development, physiology and disease. Nat. Rev. Mol. Cell Biol. 2016, 17, 240–256. [Google Scholar] [CrossRef] [PubMed]
- Ricard, J.; Salinas, J.; Garcia, L.; Liebl, D.J. EphrinB3 regulates cell proliferation and survival in adult neurogenesis. Mol. Cell. Neurosci. 2006, 31, 713–722. [Google Scholar] [CrossRef] [PubMed]
- García-Ceca, J.; Alfaro, D.; Montero-Herradón, S.; Zapata, A.G. Eph/ephrinB signalling is involved in the survival of thymic epithelial cells. Immunol. Cell Biol. 2013, 91, 130–138. [Google Scholar] [CrossRef]
- Alfaro, D.; Zapata, A.G. Eph/Ephrin-mediated stimulation of human bone marrow mesenchymal stromal cells correlates with changes in cell adherence and increased cell death. Stem Cell Res. Ther. 2018, 9, 172. [Google Scholar] [CrossRef] [PubMed]
- White, B.E.P.; Getsios, S. Eph receptor and ephrin function in breast, gut, and skin epithelia. Cell Adhes. Migr. 2014, 8, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, D.G. Regulation of cell differentiation by Eph receptor and ephrin signaling. Cell Adhes. Migr. 2014, 8, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Palmer, A.; Klein, R. Multiple Roles of Ephrins in Morphogenesis, Neuronal Networking, and Brain Function. Genes Dev. 2003, 17, 1429–1450. [Google Scholar] [CrossRef]
- Surawska, H.; Ma, P.C.; Salgia, R. The role of ephrins and Eph receptors in cancer. Cytokine Growth Factor Rev. 2004, 15, 419–433. [Google Scholar] [CrossRef]
- Ireton, R.; Chen, J. EphA2 Receptor Tyrosine Kinase as a Promising Target for Cancer Therapeutics. Curr Cancer Drug Targets 2005, 5, 149–157. [Google Scholar] [CrossRef]
- Charles, M.S.K.; Landen, N.; Sood, A.K. EphA2 as a target for ovarian cancer therapy. Expert Opin. Ther. Targets 2005, 9, 1179–1187. [Google Scholar]
- Wykosky, J.; Debinski, W. The EphA2 Receptor and EphrinA1 Ligand in Solid Tumors: Function and Therapeutic Targeting. Mol. Cancer Res. 2008, 6, 1795–1806. [Google Scholar] [CrossRef] [PubMed]
- Nikas, I.; Giaginis, C.; Petrouska, K.; Alexandrou, P.; Michail, A.; Sarantis, P.; Tsourouflis, G.; Danas, E.; Pergaris, A.; Politis, P.K.; et al. EPHA2, EPHA4, and EPHA7 Expression in Triple-Negative Breast Cancer. Diagnostics 2022, 12, 366. [Google Scholar] [CrossRef] [PubMed]
- Karidis, N.P.; Giaginis, C.; Tsouroufis, G.; Alexandrou, P.; Delladetsima, I.; Theocharis, S. Eph-A2 and Eph-A4 expression in human benign and malignant thyroid lesions: An immunohistochemical study. Med. Sci. Monit. 2011, 17, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Noren, N.K.; Pasquale, E.B. Paradoxes of the EphB4 receptor in cancer. Cancer Res. 2007, 67, 3994–3997. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, G.; Brantley-Sieders, D.M.; Vaught, D.; Yu, J.; Xie, L.; Wells, S.; Jackson, D.; Muraoka-Cook, R.; Arteaga, C.; Chen, J. Elevation of Receptor Tyrosine Kinase EphA2 Mediates Resistance to Trastuzumab Therapy. Cancer Res. 2010, 70, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.R.; Masood, R.; Spannuth, W.A.; Singh, J.; Scehnet, J.; Kleiber, G.; Jennings, N.; Deavers, M.; Krasnoperov, V.; Dubeau, L. Preferential induction of EphB4 over EphB2 and its implication in colorectal cancer progression. Cancer Res. 2009, 69, 3736–3745. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.R.; Scehnet, J.S.; Ley, E.J.; Singh, J.; Krasnoperov, V.; Liu, R.; Manchanda, P.K.; Ladner, R.D.; Hawes, D.; Weaver, F.A. The receptor tyrosine kinase EphB4 is overexpressed in ovarian cancer, provides survival signals and predicts poor outcome. Br. J. Cancer 2007, 96, 1083–1091. [Google Scholar] [CrossRef]
- Giaginis, C.; Alexandrou, P.; Poulaki, E.; Delladetsima, I.; Troungos, C.; Patsouris, E.; Theocharis, S. Clinical Significance of EphB4 and EphB6 Expression in Human Malignant and Benign Thyroid Lesions. Pathol. Oncol. Res. 2016, 22, 269–275. [Google Scholar] [CrossRef]
- Pergaris, A.; Danas, E.; Goutas, D.; Sykaras, A.G.; Soranidis, A.; Theocharis, S. The Clinical Impact of the EPH/Ephrin System in Cancer: Unwinding the Thread. Int. J. Mol. Sci. 2021, 22, 8412. [Google Scholar] [CrossRef]
- Bakst, R.L.; Wong, R.J. Mechanisms of Perineural Invasion. J. Neurol. Surg. Part B Skull Base 2016, 77, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Ravasio, A.; Myaing, M.Z.; Chia, S.; Arora, A.; Sathe, A.; Cao, E.Y.; Bertocchi, C.; Sharma, A.; Arasi, B.; Chung, V.Y.; et al. Single-cell analysis of EphA clustering phenotypes to probe cancer cell heterogeneity. Commun. Biol. 2020, 3, 429. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Li, M.; Lei, Y.; Lu, S.; Wang, S.; Liu, Z.; Wang, C.; Zhao, Y.; Wang, A.; Bi, C.; et al. An integrated analysis of single-cell and bulk transcriptomics reveals EFNA1 as a novel prognostic biomarker for cervical cancer. Hum. Cell 2022, 35, 705–720. [Google Scholar] [CrossRef] [PubMed]
- Xi, H.Q.; Wu, X.S.; Wei, B.; Chen, L. Eph receptors and ephrins as targets for cancer therapy. J. Cell. Mol. Med. 2012, 16, 2894–2909. [Google Scholar] [CrossRef] [PubMed]
- Cancer Invasion and Metastasis: Molecular and Cellular Perspective, Madame Curie Bioscience Database. Available online: https://www.ncbi.nlm.nih.gov/books/NBK5974/ (accessed on 20 February 2024).
- Cramer, K.S.; Miko, I.J. Eph-ephrin signaling in nervous system development. F1000Research 2016, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Steinecke, A.; Gampe, C.; Zimmer, G.; Rudolph, J.; Bolz, J. EphA/ephrin A reverse signaling promotes the migration of cortical interneurons from the medial ganglionic eminence. Development 2014, 141, 460–471. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, J.; Gerstmann, K.; Zimmer, G.; Steinecke, A.; Döding, A.; Bolz, J. A dual role of EphB1/ephrin-B3 reverse signaling on migrating striatal and cortical neurons originating in the preoptic area: Should I stay or go away? Front. Cell. Neurosci. 2014, 8, 185. [Google Scholar] [CrossRef] [PubMed]
- Laussu, J.; Khuong, A.; Gautrais, J.; Davy, A. Beyond boundaries—Eph: Ephrin signaling in neurogenesis. Cell Adhes. Migr. 2014, 8, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Khodosevich, K.; Watanabe, Y.; Monyer, H. EphA4 preserves postnatal and adult neural stem cells in an undifferentiated state in vivo. J. Cell Sci. 2011, 124, 1268–1279. [Google Scholar] [CrossRef]
- Jiao, J.W.; Feldheim, D.A.; Chen, D.F. Ephrins as negative regulators of adult neurogenesis in diverse regions of the central nervous system. Proc. Natl. Acad. Sci. USA 2008, 105, 8778–8783. [Google Scholar] [CrossRef]
- Hara, Y.; Nomura, T.; Yoshizaki, K.; Frisén, J.; Osumi, N. Impaired hippocampal neurogenesis and vascular formation in ephrin-A5-deficient mice. Stem Cells 2010, 28, 974–983. [Google Scholar] [CrossRef] [PubMed]
- Conover, J.C.; Doetsch, F.; Garcia-Verdugo, J.M.; Gale, N.W.; Yancopoulos, G.D.; Alvarez-Buylla, A. Disruption of Eph/Ephrin Signaling Affects Migration and Proliferation in the Adult Subventricular Zone. Nat. Neurosci. 2000, 3, 1091–1097. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y. “Medulloblastoma: Current Perspectives and Recent Advances. Brain Tumor Res. Treat. 2023, 11, 28. [Google Scholar] [CrossRef] [PubMed]
- Kool, M.; Koster, J.; Bunt, J.; Hasselt, N.E.; Lakeman, A.; van Sluis, P.; Troost, D.; Schouten-van Meeteren, N.; Caron, H.N.; Cloos, J.; et al. Integrated Genomics Identifies Five Medulloblastoma Subtypes with Distinct Genetic Profiles, Pathway Signatures and Clinicopathological Features. PLoS ONE 2008, 3, e3088. [Google Scholar] [CrossRef] [PubMed]
- Sikkema, A.H.; Den Dunnen, W.F.A.; Hulleman, E.; Van Vuurden, D.G.; Garcia-Manero, G.; Yang, H.; Scherpen, F.J.G.; Kampen, K.R.; Hoving, E.W.; Kamps, W.A.; et al. EphB2 Activity Plays a Pivotal Role in Pediatric Medulloblastoma Cell Adhesion and Invasion. Neuro-Oncology 2012, 14, 1125–1135. [Google Scholar] [CrossRef] [PubMed]
- Schlesinger, N. Abstract 1236: Ephrin Signaling in Medulloblastoma. Cancer Res. 2012, 72, 1236. [Google Scholar] [CrossRef]
- McKinney, N.; Yuan, L.; Zhang, H.; Liu, J.; Cho, Y.J.; Rushing, E.; Schniederjan, M.; MacDonald, T.J. EphrinB1 Expression Is Dysregulated and Promotes Oncogenic Signaling in Medulloblastoma. J. Neurooncol. 2015, 121, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.; Hirsch, K.; Baig, N.A.; Rodriguez, O.; Timofeeva, O.; Kavanagh, K.; Lee, Y.C.; Wang, X.J.; Albanese, C.; Karam, S.D. Effects of Altered Ephrin-A5 and EphA4/EphA7 Expression on Tumor Growth in a Medulloblastoma Mouse Model. J. Hematol. Oncol. 2015, 8, 105. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.; Baig, N.A.; Timofeeva, O.; Pasquale, E.B.; Hirsch, K.; MacDonald, T.J.; Dritschilo, A.; Lee, Y.C.; Henkemeyer, M.; Rood, B.; et al. Knockdown of Ephb1 Receptor Decreases Medulloblastoma Cell Growth and Migration and Increases Cellular Radiosensitization. Oncotarget 2015, 6, 8929–8946. [Google Scholar] [CrossRef]
- Nakada, M.; Hayashi, Y.; Hamada, J.I. Role of Eph/ephrin tyrosine kinase in malignant glioma. Neuro-Oncology 2011, 13, 1163–1170. [Google Scholar] [CrossRef]
- Okada, H.; Low, K.L.; Kohanbash, G.; McDonald, H.A.; Hamilton, R.L.; Pollack, I.F. Expression of glioma-associated antigens in pediatric brain stem and non-brain stem gliomas. J. Neurooncol. 2008, 88, 245–250. [Google Scholar] [CrossRef]
- Liu, F.; Park, P.J.; Lai, W.; Maher, E.; Chakravarti, A.; Durso, L.; Jiang, X.; Yu, Y.; Brosius, A.; Thomas, M.; et al. A Genome-Wide Screen Reveals Functional Gene Clusters in the Cancer Genome and Identifies EphA2 as a Mitogen in Glioblastoma. Cancer Res. 2006, 66, 10815–10823. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.F.; Fokas, E.; Juricko, J.; You, A.; Rose, F.; Pagenstecher, A.; Engenhart-Cabillic, R.; An, H.X. Increased Expression of EphA7 Correlates with Adverse Outcome in Primary and Recurrent Glioblastoma Multiforme Patients. BMC Cancer 2008, 8, 79. [Google Scholar] [CrossRef]
- Miao, H.; Li, D.Q.; Mukherjee, A.; Guo, H.; Petty, A.; Cutter, J.; Basilion, J.P.; Sedor, J.; Wu, J.; Danielpour, D.; et al. EphA2 mediates ligand-dependent inhibition and ligand-independent promotion of cell migration and invasion via a reciprocal regulatory loop with Akt. Cancer Cell 2009, 16, 9–20. [Google Scholar] [CrossRef]
- Fukai, J.; Yokote, H.; Yamanaka, R.; Arao, T.; Nishio, K.; Itakura, T. EphA4 promotes cell proliferation and migration through a novel EphA4-FGFR1 signaling pathway in the human glioma U251 cell line. Mol. Cancer Ther. 2008, 7, 2768–2778. [Google Scholar] [CrossRef] [PubMed]
- Nakada, M.; Niska, J.A.; Miyamori, H.; McDonough, W.S.; Wu, J.; Sato, H.; Berens, M.E. The phosphorylation of EphB2 receptor regulates migration and invasion of human glioma cells. Cancer Res. 2004, 64, 3179–3185. [Google Scholar] [CrossRef]
- Nakada, M.; Niska, J.A.; Tran, N.L.; McDonough, W.S.; Berens, M.E. EphB2/R-Ras signaling regulates glioma cell adhesion, growth, and invasion. Am. J. Pathol. 2005, 2, 565–576. [Google Scholar] [CrossRef]
- Nakada, M.; Anderson, E.M.; Demuth, T.; Nakada, S.; Reavie, L.B.; Drake, K.L.; Hoelzinger, D.B.; Berens, M.E. The phosphorylation of ephrin-B2 ligand promotes glioma cell migration and invasion. Int. J. Cancer 2010, 126, 1155–1165. [Google Scholar] [CrossRef] [PubMed]
- Sawamiphak, S.; Seidel, S.; Essmann, C.L.; Wilkinson, G.A.; Pitulescu, M.E.; Acker, T.; Acker-Palmer, A. Ephrin-B2 regulates VEGFR2 function in developmental and tumour angiogenesis. Nature 2010, 465, 487–491. [Google Scholar] [CrossRef]
- Nakada, M.; Drake, K.L.; Nakada, S.; Niska, J.A.; Berens, M.E. Ephrin-B3 ligand promotes glioma invasion through activation of Rac1. Cancer Res. 2006, 66, 8492–8500. [Google Scholar] [CrossRef]
- Hoelzinger, D.B.; Mariani, L.; Weis, J.; Woyke, T.; Berens, T.J.; McDonough, W.S.; Sloan, A.; Coons, S.W.; Berens, M.E. Gene expression profile of glioblastoma multiforme invasive phenotype points to new therapeutic targets. Neoplasia 2005, 7, 7–16. [Google Scholar] [CrossRef]
- Li, J.-J.; Liu, D.-P.; Liu, G.-T.; Xie, D. EphrinA5 acts as a tumor suppressor in glioma by negative regulation of epidermal growth factor receptor. Oncogene 2009, 28, 1759–1768. [Google Scholar] [CrossRef]
- Ferluga, S.; Debinski, W. Ephs and Ephrins in malignant gliomas. Growth Factors 2014, 32, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Monclús, S.; López-Alemany, R.; Almacellas-Rabaiget, O.; Herrero-Martín, D.; Huertas-Martinez, J.; Lagares-Tena, L.; Alba-Pavón, P.; Hontecillas-Prieto, L.; Mora, J.; de Álava, E.; et al. EphA2 receptor is a key player in the metastatic onset of Ewing sarcoma. Int. J. Cancer 2018, 143, 1188–1201. [Google Scholar] [CrossRef] [PubMed]
- Miklja, Z.; Yadav, V.N.; Cartaxo, R.T.; Siada, R.; Thomas, C.C.; Cummings, J.R.; Mullan, B.; Stallard, S.; Paul, A.; Bruzek, A.K.; et al. Everolimus improves the efficacy of dasatinib in PDGFRα-driven glioma. J. Clin. Investig. 2020, 130, 5313–5325. [Google Scholar] [CrossRef] [PubMed]
- Study of Dasatinib in Combination With Everolimus for Children and Young Adults With Gliomas Harboring Platelet-Derived Growth Factor Receptor (PDGFR) Alterations. Available online: https://clinicaltrials.gov/study/NCT03352427 (accessed on 20 February 2024).
- Kuijper, S.; Turner, C.J.; Adams, R.H. Regulation of Angiogenesis by Eph–Ephrin Interactions. Trends Cardiovasc. Med. 2007, 17, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Kertesz, N.; Krasnoperov, V.; Reddy, R.; Leshanski, L.; Kumar, S.R.; Zozulya, S.; Gill, P.S. The Soluble Extracellular Domain of EphB4 (SEphB4) Antagonizes EphB4-EphrinB2 Interaction, Modulates Angiogenesis, and Inhibits Tumor Growth. Blood 2006, 107, 2330–2338. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.; Bukkapatnam, S.; Van Court, B.; Phan, A.; Oweida, A.; Gadwa, J.; Mueller, A.C.; Piper, M.; Darragh, L.; Nguyen, D.; et al. The Effects of EphrinB2 Signaling on Proliferation and Invasion in Glioblastoma Multiforme. Mol. Carcinog. 2020, 59, 1064–1075. [Google Scholar] [CrossRef] [PubMed]
- Nagaraja, S.; Vitanza, N.A.; Woo, P.J.; Taylor, K.R.; Liu, F.; Zhang, L.; Li, M.; Meng, W.; Ponnuswami, A.; Sun, W.; et al. Transcriptional Dependencies in Diffuse Intrinsic Pontine Glioma. Cancer Cell 2017, 31, 635–652.e6. [Google Scholar] [CrossRef]
- Wyss, J.; Frank, N.A.; Soleman, J.; Scheinemann, K. Novel Pharmacological Treatment Options in Pediatric Glioblastoma—A Systematic Review. Cancers 2022, 14, 2814. [Google Scholar] [CrossRef]
- Jünger, S.T.; Timmermann, B.; Pietsch, T. Pediatric ependymoma: An overview of a complex disease. Child’s Nerv. Syst. 2021, 37, 2451–2463. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.D.; Poppleton, H.; Fuller, C.; Su, X.; Liu, Y.; Jensen, P.; Magdaleno, S.; Dalton, J.; Calabrese, C.; Board, J.; et al. Radial glia cells are candidate stem cells of ependymoma. Cancer Cell 2005, 8, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Rossi, N.; Priddy, S.; Pierson, C.R.; Studebaker, A.W.; Johnson, R.A. EphB2 activation is required for ependymoma development as well as inhibits differentiation and promotes proliferation of the transformed cell. Sci. Rep. 2015, 5, 9248. [Google Scholar] [CrossRef] [PubMed]
- Gump, J.M.; Donson, A.M.; Birks, D.K.; Amani, V.M.; Rao, K.K.; Griesinger, A.M.; Kleinschmidt-DeMasters, B.K.; Johnston, J.M.; Anderson, R.C.; Rosenfeld, A.; et al. Identification of targets for rational pharmacological therapy in childhood craniopharyngioma. Acta Neuropathol. Commun. 2015, 3, 30. [Google Scholar] [CrossRef] [PubMed]
- Angus, S.P.; Oblinger, J.L.; Stuhlmiller, T.J.; DeSouza, P.A.; Beauchamp, R.L.; Witt, L.; Chen, X.; Jordan, J.T.; Gilbert, T.S.K.; Stemmer-Rachamimov, A.; et al. Children’s Tumor Foundation Synodos for NF2 Consortium. EPH receptor signaling as a novel therapeutic target in NF2-deficient meningioma. Neuro-Oncology 2018, 20, 1185–1196. [Google Scholar] [CrossRef] [PubMed]
- Parrinello, S.; Napoli, I.; Ribeiro, S.; Wingfield Digby, P.; Fedorova, M.; Parkinson, D.B.; Doddrell, R.D.; Nakayama, M.; Adams, R.H.; Lloyd, A.C. EphB signaling directs peripheral nerve regeneration through Sox2-dependent Schwann cell Sorting. Cell 2010, 143, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Zafar, A.; Wang, W.; Liu, G.; Wang, X.; Xian, W.; McKeon, F.; Foster, J.; Zhou, J.; Zhang, R. Molecular targeting therapies for neuroblastoma: Progress and challenges. Med. Res. Rev. 2021, 41, 961–1021. [Google Scholar] [CrossRef]
- Tang, X.X.; Zhao, H.; Robinson, M.E.; Cohen, B.; Cnaan, A.; London, W.; Cohn, S.L.; Cheung, N.K.; Brodeur, G.M.; Evans, A.E.; et al. Implications of EPHB6, EFNB2, and EFNB3 expressions in human neuroblastoma. Proc. Natl. Acad. Sci. USA 2000, 97, 10936–10941. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.X.; Evans, A.E.; Zhao, H.; Cnaan, A.; Brodeur, G.M.; Ikegaki, N. Association among EPHB2, TrkA, and MYCN expression in low-stage neuroblastomas. Med. Pediatr. Oncol. 2001, 36, 80–82. [Google Scholar] [CrossRef]
- Kung, B.; Zhao, H.; Hicks, S.L.; Tang, X.X.; Ikegaki, N. Biological significance of EPHA2 expression in neuroblastoma. Int. J. Oncol. 2009, 35, 845–850. [Google Scholar]
- Li, Y.; Ohira, M.; Zhou, Y.; Xiong, T.; Luo, W.; Yang, C.; Li, X.; Gao, Z.; Zhou, R.; Nakamura, Y.; et al. Genomic analysis-integrated whole-exome sequencing of neuroblastomas identifies genetic mutations in axon guidance pathway. Oncotarget 2017, 8, 56684–56697. [Google Scholar] [CrossRef]
- Chen, Q.; Deng, R.; Zhao, X.; Yuan, H.; Zhang, H.; Dou, J.; Chen, R.; Jin, H.; Wang, Y.; Huang, J.; et al. Sumoylation of EphB1 Suppresses Neuroblastoma Tumorigenesis via Inhibiting PKCγ Activation. Cell. Physiol. Biochem. 2018, 45, 2283–2292. [Google Scholar] [CrossRef] [PubMed]
- Andolfo, I.; Lasorsa, V.A.; Manna, F.; Rosato, B.E.; Formicola, D.; Iolascon, A.; Capasso, M. Kinome multigenic panel identified novel druggable EPHB4-V871I somatic variant in high-risk neuroblastoma. J. Cell. Mol. Med. 2020, 24, 6459–6471. [Google Scholar] [CrossRef]
- Matsuo, K.; Otaki, N. “Bone cell interactions through Eph/ephrin. Cell Adh. Migr. 2012, 6, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Lamont, R.E.; Ryan, E.; Childs, S. MAPping out arteries and veins. Sci. STKE Signal Transduct. Knowl. Environ. 2006, 355, e39. [Google Scholar] [CrossRef]
- Wang, Y.; Nakayama, M.; Pitulescu, M.E.; Schmidt, T.S.; Bochenek, M.L.; Sakakibara, A.; Adams, S.; Davy, A.; Deutsch, U.; Lüthi, U.; et al. Ephrin-B2 controls VEGF-induced angiogenesis and lymphangiogenesis. Nature 2010, 465, 483–486. [Google Scholar] [CrossRef]
- Zhao, C.; Irie, N.; Takada, Y.; Shimoda, K.; Miyamoto, T.; Nishiwaki, T.; Suda, T.; Matsuo, K. Bidirectional EphrinB2-EphB4 Signaling Controls Bone Homeostasis. Cell Metab. 2006, 4, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.F.; Fernandez-Pineda, I.; Gosain, A. Pediatric Sarcomas. Surg. Clin. N. Am. 2016, 96, 1107–1125. [Google Scholar] [CrossRef]
- Sáinz-Jaspeado, M.; Huertas-Martinez, J.; Lagares-Tena, L.; Martin Liberal, J.; Mateo-Lozano, S.; de Alava, E.; de Torres, C.; Mora, J.; Del Muro, X.G.; Tirado, O.M. EphA2-Induced Angiogenesis in Ewing Sarcoma Cells Works through bFGF Production and Is Dependent on Caveolin-1. PLoS ONE 2013, 8, e71449. [Google Scholar] [CrossRef]
- Fritsche-Guenther, R.; Noske, A.; Ungethüm, U.; Kuban, R.J.; Schlag, P.M.; Tunn, P.U.; Karle, J.; Krenn, V.; Dietel, M.; Sers, C. De novo expression of EphA2 in osteosarcoma modulates activation of the mitogenic signalling pathway. Histopathology 2010, 57, 836–850. [Google Scholar] [CrossRef]
- Yu, L.; Xia, K.; Gao, T.; Chen, J.; Zhang, Z.; Sun, X.; Simões, B.M.; Eyre, R.; Fan, Z.; Guo, W.; et al. The Notch Pathway Promotes Osteosarcoma Progression through Activation of Ephrin Reverse Signaling. Mol. Cancer Res. 2019, 17, 2383–2394. [Google Scholar] [CrossRef] [PubMed]
- Aggerholm-Pedersen, N.; Rossen, P.; Rose, H.; Safwat, A. Pazopanib in the Treatment of Bone Sarcomas: Clinical Experience. Transl. Oncol. 2020, 13, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Frankel, P.; Ruel, C.; Uche, A.; Choy, E.; Okuno, S.; Somiah, N.; Chow, W.A. Pazopanib in Patients with Osteosarcoma Metastatic to the Lung: Phase 2 Study Results and the Lessons for Tumor Measurement. J. Oncol. 2022, 2022, 3691025. [Google Scholar] [CrossRef] [PubMed]
- Chiabotto, G.; Grignani, G.; Todorovic, M.; Martin, V.; Centomo, M.L.; Prola, E.; Giordano, G.; Merlini, A.; Miglio, U.; Berrino, E.; et al. Pazopanib and trametinib as a synergistic strategy against osteosarcoma: Preclinical activity and molecular insights. Cancers 2020, 12, 1519. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.; Cai, Q.; Zhu, X.; Xu, M. Down-regulation of hcp5 inhibits cell proliferation, migration, and invasion through regulating epha7 by competitively binding mir-101 in osteosarcoma. Braz. J. Med. Biol. Res. 2021, 54, e9161. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yan, L.; Liu, Y.; Xian, W.; Wang, L.; Ding, X. MicroRNA-448 suppresses osteosarcoma cell proliferation and invasion through targeting EPHA7. PLoS ONE 2017, 12, e0175553. [Google Scholar] [CrossRef] [PubMed]
- Abdou, A.G.; Abd El-Wahed, M.M.; Asaad, N.Y.; Samaka, R.M.; Abdallaha, R. Ephrin A4 expression in osteosarcoma, impact on prognosis, and patient outcome. Indian J. Cancer 2010, 47, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Berardi, A.C.; Marsilio, S.; Rofani, C.; Salvucci, O.; Altavista, P.; Perla, F.M.; Diomedi-Camassei, F.; Uccini, S.; Kokai, G.; Landuzzi, L.; et al. Up-regulation of EphB and ephrin-B expression in rhabdomyosarcoma. Anticancer Res. 2008, 28, 763–769. [Google Scholar]
- Randolph, M.E.; Cleary, M.M.; Bajwa, Z.; Svalina, M.N.; Young, M.C.; Mansoor, A.; Kaur, P.; Bult, C.J.; Goros, M.W.; Michalek, J.E.; et al. EphB4/EphrinB2 therapeutics in Rhabdomyosarcoma. PLoS ONE 2017, 12, e0183161. [Google Scholar] [CrossRef]
- Aslam, M.I.; Abraham, J.; Mansoor, A.; Druker, B.J.; Tyner, J.W.; Keller, C. PDGFRβ reverses EphB4 signaling in alveolar rhabdomyosarcoma. Proc. Natl. Acad. Sci. USA 2014, 111, 6383–6388. [Google Scholar] [CrossRef]
- Bai, Y.; Li, J.; Fang, B.; Edwards, A.; Zhang, G.; Bui, M.; Eschrich, S.; Altiok, S.; Koomen, J.; Haura, E.B. Phosphoproteomics identifies driver tyrosine kinases in sarcoma cell lines and tumors. Cancer Res. 2012, 72, 2501–2511. [Google Scholar] [CrossRef] [PubMed]
- Weiss, A.-C.; Kispert, A. Eph/ephrin signaling in the kidney and lower urinary tract. Pediatr. Nephrol. 2016, 31, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Pater, L.; Melchior, P.; Rübe, C.; Cooper, B.T.; McAleer, M.F.; Kalapurakal, J.A.; Paulino, A.C. Wilms tumor. Pediatr. Blood Cancer 2021, 68, e28257. [Google Scholar] [CrossRef] [PubMed]
- Chetcuti, A.; Aktas, S.; Mackie, N.; Ulger, C.; Toruner, G.; Alkan, M.; Catchpoole, D. Expression profiling reveals MSX1 and EphB2 expression correlates with the invasion capacity of Wilms tumors. Pediatr. Blood Cancer 2011, 57, 950–957. [Google Scholar] [CrossRef]
- Bennett, B.D.; Wang, Z.; Kuang, W.J.; Wang, A.; Groopman, J.E.; Goeddel, D.V.; Scadden, D.T. Cloning and characterization of HTK, a novel transmembrane tyrosine kinase of the EPH subfamily. J. Biol. Chem. 1994, 269, 14211–14218. [Google Scholar] [CrossRef]
- Stergiou, I.E.; Papadakos, S.P.; Karyda, A.; Tsitsilonis, O.E.; Dimopoulos, M.A.; Theocharis, S. EPH/Ephrin Signaling in Normal Hematopoiesis and Hematologic Malignancies: Deciphering Their Intricate Role and Unraveling Possible New Therapeutic Targets. Cancers 2023, 15, 3963. [Google Scholar] [CrossRef] [PubMed]
- Lato, M.W.; Przysucha, A.; Grosman, S.; Zawitkowska, J.; Lejman, M. The new therapeutic strategies in pediatric t-cell acute lymphoblastic leukemia. Int. J. Mol. Sci. 2021, 22, 4502. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, H.; Chen, X.; Mai, H.; Li, C.; Wen, F. Aberrant EPHB4 gene methylation and childhood acute lymphoblastic leukemia. Oncol. Lett. 2017, 14, 4433–4440. [Google Scholar] [CrossRef] [PubMed]
- El Zawily, A.; McEwen, E.; Toosi, B.; Vizeacoumar, F.S.; Freywald, T.; Vizeacoumar, F.J.; Freywald, A. The EphB6 receptor is overexpressed in pediatric T cell acute lymphoblastic leukemia and increases its sensitivity to doxorubicin treatment. Sci. Rep. 2017, 7, 14767. [Google Scholar] [CrossRef]
- Colucci, M.; Trivieri, N.; Mencarelli, G.; De Santis, E.; Sansico, F.; Tamiro, F.; Visioli, A.; Barile, C.; Pracella, R.; Rossi, G.; et al. A functional role of Ephrin type-B receptor 6 (EPHB6) in T-cell acute lymphoblastic leukemia. Biomark. Res. 2023, 11, 92. [Google Scholar] [CrossRef]
- Kong, L.; Zhang, X.; Li, C.; Zhou, L. Potential therapeutic targets and small molecular drugs for pediatric B-precursor acute lymphoblastic leukemia treatment based on microarray data. Oncol. Lett. 2017, 14, 1543–1549. [Google Scholar] [CrossRef] [PubMed]
- El-Sisi, M.G.; Radwan, S.M.; Saeed, A.M.; El-Mesallamy, H.O. Serum levels of FAK and some of its effectors in adult AML: Correlation with prognostic factors and survival. Mol. Cell. Biochem. 2021, 476, 1949–1963. [Google Scholar] [CrossRef] [PubMed]
- Kampen, K.R.; Scherpen, F.J.; Garcia-Manero, G.; Yang, H.; Kaspers, G.J.; Cloos, J.; Zwaan, C.M.; van den Heuvel-Eibrink, M.M.; Kornblau, S.M.; De Bont, E.S. EphB1 suppression in acute myelogenous leukemia: Regulating the DNA damage control system. Mol. Cancer Res. 2015, 13, 982–992. [Google Scholar] [CrossRef] [PubMed]
- Giaginis, C.; Tsourouflis, G.; Zizi-Serbetzoglou, A.; Kouraklis, G.; Chatzopoulou, E.; Dimakopoulou, K.; Theocharis, S. Clinical significance of ephrin (eph)-A1, -A2, -a4, -a5 and -a7 receptors in pancreatic ductal adenocarcinoma. Pathol. Oncol. Res. 2010, 16, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Papadakos, S.P.; Stergiou, I.E.; Gkolemi, N.; Arvanitakis, K.; Theocharis, S. Unraveling the Significance of EPH/Ephrin Signaling in Liver Cancer: Insights into Tumor Progression and Therapeutic Implications. Cancers 2023, 15, 3434. [Google Scholar] [CrossRef] [PubMed]
- Papadakos, S.P.; Petrogiannopoulos, L.; Pergaris, A.; Theocharis, S. The EPH/Ephrin System in Colorectal Cancer. Int. J. Mol. Sci. 2022, 23, 2761. [Google Scholar] [CrossRef] [PubMed]
- Gajdzis, M.; Theocharis, S.; Gajdzis, P.; Cassoux, N.; Gardrat, S.; Donizy, P.; Klijanienko, J.; Kaczmarek, R. Ephrin Receptors (Eph): EphA1, EphA5, and EphA7 Expression in Uveal Melanoma-Associations with Clinical Parameters and Patient Survival. Life 2020, 10, 225. [Google Scholar] [CrossRef]
- Giaginis, C.; Tsoukalas, N.; Bournakis, E.; Alexandrou, P.; Kavantzas, N.; Patsouris, E.; Theocharis, S. Ephrin (Eph) receptor A1, A4, A5 and A7 expression in human non-small cell lung carcinoma: Associations with clinicopathological parameters, tumor proliferative capacity and patients’ survival. BMC Clin. Pathol. 2014, 14, 8. [Google Scholar] [CrossRef]
- Masaoutis, C.; Georgantzoglou, N.; Sarantis, P.; Theochari, I.; Tsoukalas, N.; Bobos, M.; Alexandrou, P.; Pergaris, A.; Rontogianni, D.; Theocharis, S. Ephrin Receptors (Ephs) Expression in Thymic Epithelial Tumors: Prognostic Implications and Future Therapeutic Approaches. Diagnostics 2021, 11, 2265. [Google Scholar] [CrossRef]
- Sarno, J.; Domizi, P.; Liu, Y.; Merchant, M.; Pedersen, C.B.; Jedoui, D.; Jager, A.; Nolan, G.P.; Gaipa, G.; Bendall, S.C.; et al. Dasatinib overcomes glucocorticoid resistance in B-cell acute lymphoblastic leukemia. Nat. Commun. 2023, 14, 2935. [Google Scholar] [CrossRef]
- Romero, D. Dabrafenib–trametinib is effective in paediatric high-grade glioma. Nat. Rev. Clin. Oncol. 2023, 20, 734. [Google Scholar] [CrossRef] [PubMed]
- Duffaud, F.; Blay, J.Y.; Le Cesne, A.; Chevreau, C.; Boudou-Rouquette, P.; Kalbacher, E.; Penel, N.; Perrin, C.; Laurence, V.; Bompas, E.; et al. Regorafenib in patients with advanced Ewing sarcoma: Results of a non-comparative, randomised, double-blind, placebo-controlled, multicentre Phase II study. Br. J. Cancer 2023, 129, 1940–1948. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Wang, W.; Fang, G. Targeting Protein-Protein Interaction by Small Molecules. Annu. Rev. Pharmacol. Toxicol. 2014, 54, 435–456. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.; Scott, A.M.; Janes, P.W. Eph Receptors in Cancer. Biomedicines 2023, 11, 315. [Google Scholar] [CrossRef] [PubMed]
- Offenhäuser, C.; Al-Ejeh, F.; Puttick, S.; Ensbey, K.S.; Bruce, Z.C.; Jamieson, P.R.; Smith, F.M.; Stringer, B.W.; Carrington, B.; Fuchs, A.V.; et al. EphA3 pay-loaded antibody therapeutics for the treatment of glioblastoma. Cancers 2018, 10, 519. [Google Scholar] [CrossRef] [PubMed]
- Mao, W.; Luis, E.; Ross, S.; Silva, J.; Tan, C.; Crowley, C.; Chui, C.; Franz, G.; Senter, P.; Koeppen, H.; et al. EphB2 as a Therapeutic Antibody Drug Target for the Treatment of Colorectal Cancer. Cancer Res. 2004, 64, 781–788. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Suo, Z.; Risberg, B.; Karlsson, M.G.; Villman, K.; Nesland, J.M. Expression of Ephb2 and Ephb4 in breast carcinoma. Pathol. Oncol. Res. 2004, 10, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Palath, V.; Vekhande, R.; Baer, M.; Ching, W.; Tomasevic, N.; Martinez, D.; Yi, C.; Bhaskar, S.; Luehrsen, K.; Williams, J.; et al. A Recombinant Antibody to EphA3 with Pro-Apoptotic and Enhanced ADCC Activity Against Various Hematologic Malignancies Shows Selective Inhibition of Colony Formation from Long-Term Culture-Initiating Cells (LTC-ICs) In Primary Leukemia Samples. Blood 2010, 116, 2897. [Google Scholar] [CrossRef]
- Annunziata, C.M.; Kohn, E.C.; LoRusso, P.; Houston, N.D.; Coleman, R.L.; Buzoianu, M.; Robbie, G.; Lechleider, R. Phase 1, open-label study of MEDI-547 in patients with relapsed or refractory solid tumors. Investig. New Drugs 2013, 31, 77–84. [Google Scholar] [CrossRef]
- Duggineni, S.; Mitra, S.; Lamberto, I.; Han, X.; Xu, Y.; An, J.; Pasquale, E.B.; Huang, Z. Design and Synthesis of Potent Bivalent Peptide Agonists Targeting the EphA2 Receptor. ACS Med. Chem. Lett. 2013, 4, 344–348. [Google Scholar] [CrossRef]
- Kantarjian, H.; Jabbour, E.; Grimley, J.; Kirkpatrick, P. Dasatinib. Nat. Rev. Drug Discov. 2006, 5, 717–718. [Google Scholar] [CrossRef] [PubMed]
- Fondevila, F.; Méndez-Blanco, C.; Fernández-Palanca, P.; González-Gallego, J.; Mauriz, J.L. Anti-tumoral activity of single and combined regorafenib treatments in preclinical models of liver and gastrointestinal cancers. Exp. Mol. Med. 2019, 51, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.M.; Real, A.M.; Marsiglia, W.M.; Chow, A.; Duffy, M.E.; Yerabolu, J.R.; Scopton, A.P.; Dar, A.C. Structural basis for the action of the drug trametinib at KSR-bound MEK. Nature 2020, 588, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.T.J.; Jones, R.L.; Huang, P.H. Pazopanib in advanced soft tissue sarcomas. Signal Transduct. Target. Ther. 2019, 4, 16. [Google Scholar] [CrossRef]
- Singh, A.; Patel, V.K.; Jain, D.K.; Patel, P.; Rajak, H. Panobinostat as Pan-deacetylase Inhibitor for the Treatment of Pancreatic Cancer: Recent Progress and Future Prospects. Oncol. Ther. 2016, 4, 73–89. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EPH/Ephrin Member | Embryonic Developmental Function | Postnatal/Adult Normal (Homeostatic) Function | Tumor Type | Expression Pattern in Tumor | Tumorigenic Function | Potential Targets and Targeting Effect |
|---|---|---|---|---|---|---|
| EPHA2 | CNS angiogenesis inhibition via interaction with ephrin-A1 [60] | Glioma | High-level expression in high-grade gliomas [73,74,75] | Promotion of neoplastic cell migration and proliferation [73,74,85] Ligand-mediated EPHA2 activation acts via MAPK-dependent pathway [74] Ligand-independent EPHA2 activation via phosphorylation at serine 897 by Akt [76] | Targeting with Dasatinib results in downregulation of proliferation in mouse and human cell lines [87] | |
| Medulloblastoma | High-level expression in medulloblastoma cell lines compared to normal cells [67] | |||||
| Craniopharyngioma | High-level expression compared to normal cells [97] | Targeting with Dasatinib and Regorafenib has shown promising effects in tumor inhibition [97] | ||||
| NF2-deficient meningioma | Overexpression in NF2-deficient meningioma cell lines compared to normal cells [98] | Targeting with Dasatinib results in a dose-dependent reduction in expression [98] | ||||
| Diffuse intrinsic pontine glioma | Targeting with Panobinostat results in a dose-dependent reduction in cell invasion [92] | |||||
| EPHA4 | Motility enhancement of migrating medial ganglionic eminence inter-neurons. [54] | 1. Self-renewal of radial glial cells in the neocortex via interaction with ephrin-B1 [60] 2. Postnatal and adult neural stem cell preservation in an undifferentiated state in vivo via interaction with ephrin-A5 [61] 3. Varying levels of expression in normal cerebellum cells [70] | Glioma | Inhibition of neoplastic cell migration and proliferation [85] EPHA4 promotes FGF2-mediated cell proliferation and migration and increased MAPK and Akt phosphorylation stimulated by FGF2 [77] EPHA4 overexpressing cells show increased active forms of Rac1 and Cdc42 [77] EPHA4 forms a heteroreceptor complex with FGFR1 inducing FGFR1-mediated downstream signaling [77] | ||
| Diffuse intrinsic pontine glioma | Targeting with Panobinostat results in dose dose-dependent reduction in cell invasion [92] | |||||
| EPHA7 | 1. Neuronal migration and proliferation in subventricular zone (SVZ) [64] 2. Multiple-level expression in normal cerebellum cells [70] | Glioma | High-level expression in poor-prognosis gliomas [75] | Enhancement of tumorigenic effects [75] | ||
| EPHB1 | 1. Retinal ganglion cell axons direction to the ipsilateral trajectory [57] 2. Dendritic spine formation and synaptic maturation [57] 3. Neuron migration enhancement [59] | 1. Neuron antiproliferative effect [60] 2. Neuron proliferation control in SVZ [64] | Glioma | High-level expression in favorable prognosis gliomas [67] | Inhibition of neoplastic cell migration and invasion [85] | |
| Medulloblastoma | Varying levels of expression in 95% of the tumors [69] High-level expression in radiosensitive tumors [71] | Tumorigenic activity via enhancement of cell proliferation, cell migration, and upregulation of survival proteins [70] | ||||
| Diffuse intrinsic pontine glioma | Targeting with Panobinostat results in a dose-dependent reduction in cell invasion [92] | |||||
| NF2-deficient meningioma | Overexpression in all NF2-deficient meningioma cell lines [98] | Targeting with Dasatinib results in a dose-dependent reduction in expression [98] | ||||
| EPHB2 | Dendritic spine formation and synaptic maturation [57] | Early migration of subgranular zone (SGZ) progenitors [60] | Medulloblastoma | High-level expression in tumor tissue samples [66,67] | ||
| Ependymoma | Varying levels of expression in 95% of the tumors [42] | Expression required for ependymoma development [96] | ||||
| NF2-deficient meningioma | Overexpression in NF2-deficient meningioma cell lines [98] | Targeting with Dasatinib results in a dose-dependent reduction in expression [98] | ||||
| Diffuse intrinsic pontine glioma | Targeting with Panobinostat results in a dose-dependent reduction in cell invasion [92] | |||||
| EPHB3 | Dendritic spine formation and synaptic maturation [57] | Antiproliferative effect [60] | Ependymoma | High-level expression in ependymoma cells [95] | ||
| Diffuse intrinsic pontine glioma | Targeting with Panobinostat results in a dose-dependent reduction in cell invasion [92] | |||||
| EPHB4 | Medulloblastoma | High-level expression in most medulloblastoma cell lines [67] | ||||
| High-grade glioma | High-level expression [91] | Cell proliferation and invasion downregulation via EPHB4 stimulation with an ephrin-B2 fusion protein [91] | ||||
| ephrin-A2 | Negative regulator of adult neurogenesis [62] | Glioma | Expression associated with a favorable prognosis [72] | |||
| ephrin-A3 | Negative regulator of adult neurogenesis [62] | Ependymoma | High-level expression in ependymoma cell lines [95] | |||
| ephrin-A4 | Ependymoma | High-level expression in ependymoma cell lines [95] | ||||
| ephrin-A5 | 1. Postnatal and adult neural stem preservation cells in an undifferentiated state in vivo via interaction with EPHA4 [61] 2. Hippocampal neurogenesis and vascular formation in SGZ [63] | Medulloblastoma | Expression associated with tumor growth and invasive capacity [70] | |||
| ephrin-B1 | Inhibition of apical progenitors’ adhesion via GTPase Arf6 negative regulation [57] | Self-renewal of radial glial cells in the neocortex via interaction with EPHA4 [60] | Medulloblastoma | High-level expression in medulloblastoma cell lines [67] | Association with high invasive capacity via induction of EPHB2 phosphorylation and p38, ERK, and mTOR downstream signaling [66,67] | |
| ephrin-B2 | Medulloblastoma | Expression associated with high invasive capacity [68,69] | ||||
| Glioma | High-level expression [82] | Tumor invasion and angiogenesis via VEGFR2 regulation [79,81] | ||||
| ephrin-B3 | Glioma | High-level expression in aggressive glioma [81] | Tumor invasion via Rac1 activation [79,81] |
| EPH/Ephrin Member | Embryonic Developmental Function | Postnatal/Adult Normal (Homeostatic) Function | Tumor Type | Expression Pattern in Tumor | Tumorigenic Function | Potential Targets and Targeting Effect |
|---|---|---|---|---|---|---|
| EPHA2 | Neuroblastoma | Low-level expression in N-type neuroblastoma cell lines [103] | Targeting with Doxorubicin results in increased expression in neuroblastoma cells [103] | |||
| High-level expression in S-type neuroblastoma cell lines [103] | ||||||
| High-level expression in neuroblastoma cells associated with better prognosis [103] | ||||||
| EPHB1 | Neuroblastoma | Tumorigenesis via PKCγ activation [105] | ||||
| EPHB2 | Schwann cells guidance in collective migration, forming multicellular cords that facilitate axon guidance across the injury site [89] | Neuroblastoma | Expression in low-stage tumors correlated with MYCN expression [102] | |||
| EPHB4 | Neuroblastoma | Expression in neuroblastoma cells [106] | EPHB4-V871I variant associated with increased proliferation, migration, and invasion properties via the ERK1-2 pathway and VEGF, c-RAF, and CDK4 target genes [106] | Pharmaceutical targeting inhibits kinase activity obstructing tumor progression in neuroblastoma cell lines [106] | ||
| EPHB6 | Neuroblastoma | Expression in tumor specimens associated with a favorable prognosis [101] | Tumorigenesis via involvement in axon guidance pathways [104] | |||
| ephrin-B2 | Neuroblastoma | Expression in tumor specimens associated with a favorable prognosis [101] | ||||
| ephrin-B3 | Neurons migration [59] | Neuroblastoma | Expression in tumor specimens associated with a favorable prognosis [101] |
| Eph/Ephrin Member | Embryonic Developmental Function | Postnatal/Adult Normal (Homeostatic) Function | Tumor Type | Expression Pattern in Tumor | Tumorigenic Function | Potential Targets and Targeting Effect |
|---|---|---|---|---|---|---|
| EPHA2 | Ewing sarcoma | Overexpression in Ewing sarcoma compared to normal tissue, especially in males [114] | 1. EPHA2 and CAV1 interaction induces AKT-mediated bFGF production, promoting endothelial cell migration angiogenetic activity [112] 2. EPHA2 phosphorylation correlated with the capacity of Ewing sarcoma cell migration [86] | |||
| Osteosarcoma | High-level expression in osteosarcoma, especially in males and in tumors with high invasive capacity [113,114] | Ephrin-A1 binding to EPHA2 induces the activation of MAPK-mediated downstream signaling pathways, stimulating osteosarcoma’s metastatic potential [113] | Pazopanib, Trametinib [115,116,117] | |||
| EPHA7 | Osteosarcoma | miR-101 binding to EPHA7 regulates EPHA7 activation—HCP5 competitive binding to miR-101 inhibits EPHA7-induced cell proliferation, migration, and invasion [118] | ||||
| EPHB1 | Rhabdomyosarcoma | Expression in rhabdomyosarcoma tumor cells [121] | ||||
| EPHB2 | Mesenchymal stem cell migration [107] | Rhabdomyosarcoma | Expression in rhabdomyosarcoma tumor cells [121] | |||
| EPHB3 | Bone growth in the embryo (especially at the calvaria suture fronts, periosteum, chondrocytes, and trabeculae of developing long bones) [107] | Rhabdomyosarcoma | Expression in rhabdomyosarcoma tumor cells [121] | |||
| EPHB4 | Angiogenesis stimulation [107] | Rhabdomyosarcoma | Expression in rhabdomyosarcoma tumor cells [121] | |||
| High-level expression in alveolar type associated with poor prognosis [123,124] | ||||||
| Low-level expression in embryonal rhabdomyosarcoma [122] | ||||||
| ephrin-A4 | Osteosarcoma | Cytoplasmic expression in osteosarcoma cells associated with poor prognosis and inferior response to chemotherapy [120] | ||||
| ephrin-B1 | Bone remodeling [107] | Rhabdomyosarcoma | Expressed in rhabdomyosarcoma tumor cells [120] | |||
| ephrin-B2 | VEGF-angiogenesis control during development [109,110] | 1. Bone remodeling [97] 2. Expressed by osteoclasts [109] | Rhabdomyosarcoma | Expressed in rhabdomyosarcoma tumor cells [120] |
| Eph/Ephrin Member | Embryonic Developmental Function | Postnatal/Adult Normal (Homeostatic) Function | Tumor Type | Expression Pattern in Tumor | Tumorigenic Function | Potential Targets and Targeting Effect |
|---|---|---|---|---|---|---|
| EPHB2 | Involved in genesis, differentiation, and developmental process of renal system structures (glomerular epithelium, cloaca, and urethra) [125] | Venous and capillary maintenance and angiogenic remodeling [127] | Wilms Tumor | Increased expression in tumor samples of advanced tumor stages [127] |
| EPH/Ephrin Member | Embryonic Developmental Function | Postnatal/Adult Normal (Homeostatic) Function | Tumor Type | Expression Pattern in Neoplastic cells | Tumorigenic Function | Potential Targets and Targeting Effect |
|---|---|---|---|---|---|---|
| EPHA4 | Lymphocyte development [129] | AML | Positive expression in 36.8% of peripheral blood samples of pediatric AML cases [134] | Tumor-promoting [134] | ||
| EPHB1 | 1. HSPC maintenance 2. HSPC differentiation 3. Erythroid differentiation [129] | AML | Decreased EPHB1 phosphorylation and mRNA expression with concurrent EPHB1 promoter hypermethylation in AML bone marrow samples compared to healthy controls [136] | Tumor-suppressive-EPHB1 activation induces DDR [136] | ||
| EPHB4 | Fetal B-cell maturation | 1. Myeloid Differentiation 2. Erythroid differentiation 3. Adult B-cell maturation 4. Monocyte differentiation 5. Adhesion, migration, and extravasation of monocytes and macrophages [122] | ALL | Higher EPHB4 methylation in ALL samples compared to controls [131] | Tumor-suppressive [131] | |
| EPHB6 | Thymic T-cell development [129] | T-ALL | High-level expression associated with sensitivity to chemotherapy [132] | EPHB6-mediated preservation of Akt signaling increases chemosensitivity [132] | Doxorubicin [132] | |
| High-level expression [133] | Chemotherapy resistance via enhancement of self-proliferation processes (increased CCNB1 and KIF20A) [133] | |||||
| ephrin-B1 | 1. HSPC maintenance 2. B-cell development 3. T-cell development [129] | Β-ALL | Among downregulated DEGs enriched in the cell cycle process [132] | Tumor-suppressive [132] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chatzikalil, E.; Stergiou, I.E.; Papadakos, S.P.; Konstantinidis, I.; Theocharis, S. The Clinical Relevance of the EPH/Ephrin Signaling Pathway in Pediatric Solid and Hematologic Malignancies. Int. J. Mol. Sci. 2024, 25, 3834. https://doi.org/10.3390/ijms25073834
Chatzikalil E, Stergiou IE, Papadakos SP, Konstantinidis I, Theocharis S. The Clinical Relevance of the EPH/Ephrin Signaling Pathway in Pediatric Solid and Hematologic Malignancies. International Journal of Molecular Sciences. 2024; 25(7):3834. https://doi.org/10.3390/ijms25073834
Chicago/Turabian StyleChatzikalil, Elena, Ioanna E. Stergiou, Stavros P. Papadakos, Ippokratis Konstantinidis, and Stamatios Theocharis. 2024. "The Clinical Relevance of the EPH/Ephrin Signaling Pathway in Pediatric Solid and Hematologic Malignancies" International Journal of Molecular Sciences 25, no. 7: 3834. https://doi.org/10.3390/ijms25073834