
Does the Artificial Enhancement and Release Activity Affect the Genetic Diversity of Marbled Rockfish Sebastiscus marmoratus in Zhoushan Waters?

Abstract
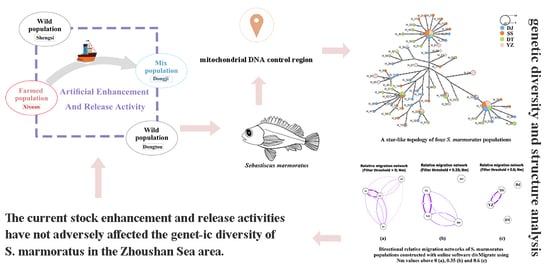
:
1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Sampling
2.2. DNA Extraction and Polymerase Chain Reaction
2.3. Data Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, M.Z.; Zeng, Y.; Ren, T.J. Study on the problems and counter-measures of fishery proliferation and release in China. China Fish. 2021, 2021, 42–45. [Google Scholar]
- Li, K.Z.; Wang, C.G.; Liang, H.Y.; Zhao, G.Q.; Li, C.Y.; Qiu, S.Y. Research on contribution rate of resources on proliferation and releasing of penaeus chinensis in the southern Shandong Peninsula. Tournal Yantai Univ. (Nat. Sci. Eng. Ed.) 2019, 32, 165–170. [Google Scholar] [CrossRef]
- Shui, B.N. Rethinking and optimization of releasing stock enhancementfor marine fishery resources: A review. J. Dalian Ocean. Univ. 2023, 38, 737–743. [Google Scholar] [CrossRef]
- Xu, K.D.; Zhou, Y.D.; Wang, Y.; Wang, W.D.; Xu, H.X.; Zhang, H.L.; Li, P.F.; Lian, J.; Chen, F.; Lu, Z.H.; et al. Effect and assessment of enhancement release of Sepiella maindroni in the northern coastal water of Zhejiang. J. Fish. Sci. China 2018, 25, 654–662. [Google Scholar] [CrossRef]
- Li, L.P.; Huang, S.L. A study on management of stock enhancement in China. J. Shanghai Ocean. Univ. 2011, 20, 765–772. [Google Scholar] [CrossRef]
- Liao, Z.; Chen, X.Y.; Hao, Y.F.; Yin, D.; Tian, P.; Zeng, S. The Application of Marking Techniques Is Used to Assess the Effects of Fishery Stocking and Release. Mod. Anim. Husb. Sci. Technol. 2024, 02, 112–114. [Google Scholar] [CrossRef]
- Laikre, L.; Schwartz, M.K.; Waples, R.S.; Ryman, N. Compromising genetic diversity in the wild: Unmonitored large-scale release of plants and animals. Trends Ecol. Evol. 2010, 25, 520–529. [Google Scholar] [CrossRef]
- Qi, J.F.; Yin, B.S.; Zhang, Q.L.; Yang, D.Z.; Xu, Z.H. Seasonal variation of the Taiwan Warm Current Water and its underlying mechanism. Chin. J. Oceanol. Limnol. 2017, 35, 1045–1060. [Google Scholar] [CrossRef]
- Wei, Q.S.; Wang, B.D.; Zhang, X.L.; Ran, X.B.; Fu, M.Z.; Sun, X.; Yu, Z.G. Contribution of the offshore detached Changjiang (Yangtze River) Diluted Water to the formation of hypoxia in summer. Sci. Total Environ. 2021, 764, 142838. [Google Scholar] [CrossRef]
- Xu, X.; Tang, W.R.; Wang, Y.B. Releasing capacity of Portunus trituberculatus enhancement in Zhoushan fishing ground and Yangtze river estuary fishing ground and their adjacent waters. South China Fish. Sci. 2019, 15, 126–132. [Google Scholar]
- Nakabo, T. Fishes of Japan: With Pictorial Keys to the Species; Tokai University Press: Tokyo, Japan, 2002; Volume 2. [Google Scholar]
- Fujita, H.; Kohda, M. Timing and sites of parturition of the viviparous scorpionfish, Sebastiscus marmoratus. Environ. Biol. Fishes 1998, 52, 225–229. [Google Scholar] [CrossRef]
- Chen, D.G.; Zhang, M.Z. Chinese Marine Fishes; Ocean University of China Press: Qindao, China, 2015. [Google Scholar]
- Sun, D.Q.; Shi, G.; Liu, X.Z.; Wang, R.X.; Xu, T.J. Genetic diversity and population structure of the marbled rockfish, Sebastiscus marmoratus, revealed by SSR markers. J. Genet. 2011, 90, e21–e24. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.H. Preliminry study on the fishries biology of Sebasticus marmoratus. Fish. Inf. Strategy 2000, 2000, 17–20. [Google Scholar]
- Wu, C.W. Biological Studies on Sebastiscus marmoratus of Zhoushan. J. Zhejiang Ocean. Univ. (Nat. Sci. Ed.) 1999, 16, 169–174. [Google Scholar]
- Wourms, J.P.; Grove, B.D.; Lombardi, J. The Maternal-Embryonic Relationship in Viviparous Fishes. In Fish Physiology; Hoar, W.S., Randall, D.J., Eds.; Academic Press: Cambridge, MA, USA, 1988; Volume 11, pp. 1–134. [Google Scholar]
- Liu, M.Z.; Yang, F.; Jiang, R.J.; Yin, R.; Wang, J.; Xiao, Y.; Ling, T.; Zhu, S.L. Trophic niche and potential carbon source of three reef-associated fishes of Zhongjieshan Islands. Ying Yong Sheng Tai Xue Bao 2023, 34, 1130–1136. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.B.; Jiang, R.J.; Lan, D.; Wang, Y.B.; Shan, X.L.; Li, P.F.; Liu, M.Z.; Li, X.F.; Yang, F.; Yin, R. Assessment of ecological carrying capacity of sebastiscus marmoratus in Zhoushan Sea Area in the Ecopath model. Oceanol. Et. Limnol. Sin. 2023, 54, 1672–1681. [Google Scholar]
- Chen, X.Y.; Xu, K.D.; Li, P.F.; Wang, H.X.; Zhou, Y.D.; Wang, M.J. Analysis of the Status of Marine Fish Proliferation and Release in Zhejiang Offshore. Ocean. Dev. Manag. 2023, 40, 128–135. [Google Scholar] [CrossRef]
- Aljanabi, S.M.; Martinez, I. Universal and rapid salt-extraction of high quality genomic DNA for PCR-based techniques. Nucleic Acids Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef]
- Han, Z.Q.; Gao, T.X.; Yanagimoto, T.; Sakurai, Y. Genetic population structure of Nibea albiflora in Yellow Sea and East China Sea. Fish. Sci. 2008, 74, 544–552. [Google Scholar] [CrossRef]
- Hall, T.A. Bioedit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Araki, H.; Schmid, C. Is hatchery stocking a help or harm? Evidence, limitations and future directions in ecological and genetic surveys. Aquaculture 2010, 308, S2–S11. [Google Scholar] [CrossRef]
- Berejikian, B.; Johnson, T.; Endicott, R.; Lee-Waltermire, J. Increases in steelhead (Oncorhynchus mykiss) redd abundance resulting from two conservation hatchery strategies in the Hamma Hamma River, Washington. Can. J. Fish. Aquat. Sci. 2008, 65, 754–764. [Google Scholar] [CrossRef]
- Agnalt, A.L. Fecundity of the European lobster (Homarus gammarus) off southwestern Norway after stock enhancement: Do cultured females produce as many eggs as wild females? ICES J. Mar. Sci. 2007, 65, 164–170. [Google Scholar] [CrossRef]
- Grant, W.; Bowen, B. Shallow population histories in deep evolutionary lineages of marine fishes: Insights from sardines and anchovies and lessons for conservation. J. Hered. 1998, 89, 415–426. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, X.M.; Li, C.H.; Zhang, H.; Yanagimoto, T.; Song, N.; Gao, T.X. Population genetic structure of Marbled Rockfish, Sebastiscus marmoratus (Cuvier, 1829), in the northwestern Pacific Ocean. ZooKeys 2019, 830, 127–144. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, G.M. Some genetic consequences of ice ages, and their role in divergence and speciation. Biol. J. Linn. Soc. 1996, 58, 247–276. [Google Scholar] [CrossRef]
- Liu, J.-X.; Gao, T.-X.; Yokogawa, K.; Zhang, Y.-P. Differential population structuring and demographic history of two closely related fish species, Japanese sea bass (Lateolabrax japonicus) and spotted sea bass (Lateolabrax maculatus) in Northwestern Pacific. Mol. Phylogenetics Evol. 2006, 39, 799–811. [Google Scholar] [CrossRef] [PubMed]
- Saavedra Sotelo, N.C.; Calderon Aguilera, L.E.; Reyes Bonilla, H.; Paz García, D.A.; López Pérez, R.A.; Cupul Magaña, A.; Cruz Barraza, J.A.; Rocha Olivares, A. Testing the genetic predictions of a biogeographical model in a dominant endemic Eastern Pacific coral (Porites panamensis) using a genetic seascape approach. Ecol. Evol. 2013, 3, 4070–4091. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Song, N.; Zhao, L.; Cai, S.; Han, Z.; Gao, T. Genomic evidence for local adaptation in the ovoviviparous marine fish Sebastiscus marmoratus with a background of population homogeneity. Sci. Rep. 2017, 7, 1562. [Google Scholar] [CrossRef]
- Jonsson, B.; Jonsson, N. Cultured Atlantic salmon in nature: A review of their ecology and interaction with wild fish. ICES J. Mar. Sci. 2006, 63, 1162–1181. [Google Scholar] [CrossRef]
- Bert, T.M.; Crawford, C.R.; Tringali, M.D.; Seyoum, S.; Galvin, J.L.; Higham, M.; Lund, C. Genetic Management of Hatchery-Based Stock Enhancement. In Genetic Management of Hatchery-Based Stock Enhancement; Bert, T.M., Ed.; Springer Netherlands: Dordrecht, The Netherlands, 2007; Volume 6, pp. 123–174. [Google Scholar]
- Leber, K.M.; Brennan, N.; Arce, S.M. Recruitment patterns of cultured juvenile pacific threadfin, Polydactylus sexfilis (Polynemidae), released along sandy marine shores in Hawaii. Bull. Mar. Sci. 1998, 62, 389–408. [Google Scholar]
- Wang, L.J.; Wu, Z.H.; Wang, Y.J.; Liu, M.X.; Song, A.H.; Liu, H.J.; You, F. Genetic Assessment of a Black Rockfish, Sebastes schlegelii, Stock Enhancement Program in Lidao Bay, China Based on Mitochondrial and Nuclear DNA Analysis. Front. Mar. Sci. 2020, 7, 94–105. [Google Scholar] [CrossRef]
- Yang, Y.Y.; Shi, Y.R.; Zhang, H.; Pan, Y.; Zu, K.W.; Gao, T.X.; Song, N. Comparative analyses of genetic diversity of broodstock, hatchery-released and captured populations of Acanthopagrus schlegelii based on the mitochondrial DNA control region sequence. Period. Ocean. Univ. China 2023, 53, 50–58. [Google Scholar] [CrossRef]
- Shuang, Y.; Na, S.; Xiumei, Z.; Yunzhou, W.; Sijie, W.; Tianxiang, G. Genetic diversity of swimming crab (Portunus trituberculatus) from four broodstock populations in stock enhancement inferred from mitochondrial control region. J. Fish. China 2014, 38, 1089–1096. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site | Abbreviation | Coordinate | Sampling Date |
|---|---|---|---|
| Shengsi | SS | 30°42′ N 122°29′ E | August 2023 |
| Dongji | DJ | 30°20′ N 122°36′ E | May 2023 |
| Xixuan | YZ | 29°53′ N 122°18′ E | November 2022 |
| Dongtou | DT | 27°50′ N 121°14′ E | August 2023 |
| Haplotype | SS | DJ | DT | YZ | Accession Numbers | Haplotype | SS | DJ | DT | YZ | Accession Numbers |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Hap_1 | 1 | PP700307 | Hap_35 | 1 | PP700341 | ||||||
| Hap_2 | 1 | PP700308 | Hap_36 | 1 | 1 | PP700342 | |||||
| Hap_3 | 1 | PP700309 | Hap_37 | 1 | PP700343 | ||||||
| Hap_4 | 4 | 1 | 3 | PP700310 | Hap_38 | 1 | PP700344 | ||||
| Hap_5 | 1 | PP700311 | Hap_39 | 1 | PP700345 | ||||||
| Hap_6 | 5 | 3 | 2 | 3 | PP700312 | Hap_40 | 1 | PP700346 | |||
| Hap_7 | 4 | 4 | 3 | 5 | PP700313 | Hap_41 | 1 | PP700347 | |||
| Hap_8 | 1 | PP700314 | Hap_42 | 1 | 1 | PP700348 | |||||
| Hap_9 | 1 | PP700315 | Hap_43 | 1 | PP700349 | ||||||
| Hap_10 | 1 | PP700316 | Hap_44 | 1 | PP700350 | ||||||
| Hap_11 | 1 | PP700317 | Hap_45 | 1 | PP700351 | ||||||
| Hap_12 | 1 | 2 | 1 | PP700318 | Hap_46 | 1 | PP700352 | ||||
| Hap_13 | 1 | PP700319 | Hap_47 | 1 | PP700353 | ||||||
| Hap_14 | 1 | PP700320 | Hap_48 | 1 | PP700354 | ||||||
| Hap_15 | 1 | 1 | PP700321 | Hap_49 | 1 | PP700355 | |||||
| Hap_16 | 1 | PP700322 | Hap_50 | 1 | PP700356 | ||||||
| Hap_17 | 1 | PP700323 | Hap_51 | 1 | PP700357 | ||||||
| Hap_18 | 1 | 1 | 2 | 1 | PP700324 | Hap_52 | 1 | PP700358 | |||
| Hap_19 | 1 | PP700325 | Hap_53 | 1 | PP700359 | ||||||
| Hap_20 | 1 | PP700326 | Hap_54 | 1 | PP700360 | ||||||
| Hap_21 | 1 | PP700327 | Hap_55 | 1 | PP700361 | ||||||
| Hap_22 | 1 | 1 | 1 | 1 | PP700328 | Hap_56 | 1 | PP700362 | |||
| Hap_23 | 1 | PP700329 | Hap_57 | 1 | PP700363 | ||||||
| Hap_24 | 1 | PP700330 | Hap_58 | 1 | PP700364 | ||||||
| Hap_25 | 1 | PP700331 | Hap_59 | 1 | PP700365 | ||||||
| Hap_26 | 1 | PP700332 | Hap_60 | 1 | PP700366 | ||||||
| Hap_27 | 1 | PP700333 | Hap_61 | 2 | PP700367 | ||||||
| Hap_28 | 1 | PP700334 | Hap_62 | 1 | PP700368 | ||||||
| Hap_29 | 1 | PP700335 | Hap_63 | 1 | PP700369 | ||||||
| Hap_30 | 1 | PP700336 | Hap_64 | 4 | PP700370 | ||||||
| Hap_31 | 1 | PP700337 | Hap_65 | 3 | PP700371 | ||||||
| Hap_32 | 1 | PP700338 | Hap_66 | 2 | PP700372 | ||||||
| Hap_33 | 1 | PP700339 | Hap_67 | 2 | PP700373 | ||||||
| Hap_34 | 1 | PP700340 | Hap_68 | 1 | PP700374 |
| Population | Sample Size | S | h | Hd | K | π | PiJC |
|---|---|---|---|---|---|---|---|
| SS | 30 | 44 | 22 | 0.961 | 9.211 | 0.020 | 0.020 |
| DJ | 32 | 44 | 26 | 0.980 | 9.698 | 0.021 | 0.022 |
| YZ | 31 | 45 | 16 | 0.944 | 11.669 | 0.025 | 0.026 |
| DT | 30 | 46 | 23 | 0.979 | 10.230 | 0.022 | 0.023 |
| Total | 123 | 73 | 68 | 0.966 | 10.240 | 0.022 |
| Population | SS | DJ | DT | YZ |
|---|---|---|---|---|
| SS | 0.021 | |||
| DJ | 0.021 | 0.022 | ||
| DT | 0.022 | 0.022 | 0.023 | |
| YZ | 0.024 | 0.024 | 0.025 | 0.026 |
| Source of Variation | d.f. | Sum of Squares | Variance Components | Percentage of Variation |
|---|---|---|---|---|
| One gene pool | ||||
| (SS, DJ, DT, YZ) | ||||
| Among populations | 3 | 17.365 | 0.022 Va | 0.44 |
| Within populations | 119 | 607.245 | 5.103 Vb | 99.56 |
| Two gene pools | ||||
| (SS, DT) (DJ, YZ) | ||||
| Among groups | 1 | 7.214 | 0.035 Va | 0.68 |
| Among populations Within groups | 2 | 10.151 | −0.001 Vb | −0.02 |
| Within populations | 119 | 607.245 | 5.103 Vc | 99.34 |
| Three gene pools | ||||
| (SS, DT) (DJ) (YZ) | ||||
| Among groups | 2 | 14.948 | 0.133 Va | 2.58 |
| Among populations Within groups | 1 | 2.417 | −0.090 Vb | −1.74 |
| Within populations | 119 | 607.245 | 5.103 Vc | 99.16 |
| Population | SS | DJ | DT | YZ |
|---|---|---|---|---|
| SS | inf | inf | 7.001 | |
| DJ | −0.006 | inf | 17.493 | |
| DT | −0.017 | −0.018 | 21.527 | |
| YZ | 0.034 * | 0.014 | 0.012 |
| Statistics | SS | DJ | DT | YZ | Mean | s.d. | |
|---|---|---|---|---|---|---|---|
| Tajima’s D test | Tajima’s D | −0.632 | −0.412 | −0.443 | 0.133 | −0.338 | 0.329 |
| p-value | 0.308 | 0.362 | 0.351 | 0.609 | 0.408 | 0.136 | |
| Fu’s FS test | FS | −0.632 | −11.579 | −7.465 | −0.008 | −6.505 | 4.799 |
| p-value | 0.013 | 0.002 | 0.007 | 0.525 | 0.137 | 0.259 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, S.; Chen, X.; Li, P.; Wang, H.; Li, J.; Xu, K.; Ye, Y. Does the Artificial Enhancement and Release Activity Affect the Genetic Diversity of Marbled Rockfish Sebastiscus marmoratus in Zhoushan Waters? Fishes 2024, 9, 161. https://doi.org/10.3390/fishes9050161
Jiang S, Chen X, Li P, Wang H, Li J, Xu K, Ye Y. Does the Artificial Enhancement and Release Activity Affect the Genetic Diversity of Marbled Rockfish Sebastiscus marmoratus in Zhoushan Waters? Fishes. 2024; 9(5):161. https://doi.org/10.3390/fishes9050161
Chicago/Turabian StyleJiang, Senping, Xinyi Chen, Pengfei Li, Haoxue Wang, Jiji Li, Kaida Xu, and Yingying Ye. 2024. "Does the Artificial Enhancement and Release Activity Affect the Genetic Diversity of Marbled Rockfish Sebastiscus marmoratus in Zhoushan Waters?" Fishes 9, no. 5: 161. https://doi.org/10.3390/fishes9050161